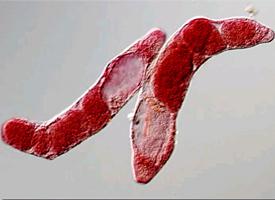
Motolice jaterní (Fasciola hepatica Linné, 1758, anglicky: common liver fluke) je kosmopolitně rozšířený parazit ze třídy Trematoda (motolice) parazitující v játrech, respektive jaterních žlučovodech ovcí, skotu a celé řady dalších savců, včetně člověka. Dospělé motolice mají dorso-ventrálně sploštělé tělo ve tvaru kopinatého listu, měří 2 - 3 cm a živí se krví definitivního hostitele. Vývojový cyklus probíhá přes mezihostitele, jimiž jsou vodní plži z čeledi plovatkovitých (Lymnaeidae). Největší význám má u ovcí a skotu. Celosvětové ztráty v zemědělství způsobené motolicí jaterní dosahují více než 2 miliard USD ročně. Infekce zvířat a lidí motolicí jaterní nebo příbuzným druhem Fasciola gigantica se označuje jako fasciolóza. Nejvíce lidí infikovaných motolicí jaterní bylo zaznamenáno v Peru, Bolívii a Ekvádoru, z Evropských států je nejvyšší prevalence u lidí ve Francii a Španělsku.
Objev vývojového cyklu
Zřejmě první písemný důkaz o motolici jaterní pochází z Francie roku 1379, kdy Jean de Brie se ve svém odborném sdělení o produkci ovčí vlny pro krále Karla V. zmiňuje i o nemoci ovcí, kterou označil jako jaterní hniloba. Nicméně ve své práci neuvedl nic o přítomnosti červů. Důkazy o přítomnosti motolic v játrech ovcí pocházejí až z první poloviny 16. století. Mělo se za to, že červi jsou důsledkem zkrmování rostlin s obsahem toxických látek. Teprve až roku 1688 italský lékař Francesco Redi popsal skutečnost, že helminti kladou vajíčka a zavrhl tak mýtus neživé podstaty vzniku červů. Není však jisté, zda popsal proces produkce vajíček právě u F. hepatica, neboť se zabýval řadou různých helmintů z různých zvířat. Redi také jako první v historii publikoval obrázek motolice jaterní izolované z jater kastrovaného berana. Podle tohoto italského vědce byly později pojmenovány larvální stádia motolic v plžích jako redie.
V roce 1698 Govert Bidloo, nizozemský profesor anatomie a zároveň osobní lékař krále Viléma III., zveřejnil, že stejný druh motolice objevil nejen u ovcí, ale i skotu, jelenů a člověka. Rovněž detekoval přítomnost vajíček uvnitř motolic. Jeho výsledky později ve svých memoárech zveřejnil zakladatel mikroskopie Anthony van Leeuwenhoek. Ten pokračoval ve výzkumech po jeho předchůdci. Leeuwenhoek se domníval, že zvířata se nakazí červy pitím vody, nicméně nedokázal vysvětlit proč nenalezl tyto organismy ve vodě (publikováno v letech 1700-1704). Další pokrok v poznání vývojového cyklu učinili v druhé polovině 18. století Johann Swammerdam (1758 – objev objev neznámých organismů v plži) a Otto Müller (1770 – objev plovoucích organismů ve vodě, které pojmenoval jako cerkarie). Müller se chybně domníval, že se jedná o Protozoa nebo Rotifera.[6] Spojitost cerkarií a larvalních stadií v plži s dospělými motolicemi jakožto parazity obratlovců zůstala neodhalena až do druhé poloviny 19. století. Mezitím bylo publikováno několik dílčích objevů. Christian Nietzsch (1807 a 1819) referoval o encystaci cerkarií na vegetaci (1807), rovněž upozornil na morfologickou podobnost cerkarií a motolic obratlovců. Mehlis (1837) pozoroval líhnutí miracidií z vajíček motolic a domníval se, že důvodem jejich intezivního pohybu ve vodě je snaha najít nějakého hostitele. Jediným vědcem této doby, který zastával teorii o existenci mezihostitele ve vývoji parazitárních ploštěnců, byl Peter Abildgaard. Ten roku 1790 podal důkaz o tom, že larvální vývoj tasemnice Diphyllobothrium spp. probíhá přes rybího mezihostitele. Jeho výsledky nenašly velký ohlas a byly na dlouhou dobu zapomenuty. Teprve v roce 1881 byl podán důkaz, že vývojový cyklus motolice jaterní probíhá přes plže Galba truncatula. Tento přelomový objev v biologii F. hepatica, potažmo všech motolic, byl učiněn současně, nezávisle na sobě Algeronem Thomasem ve Spojeném Království a známým zoologem Rudolfem Leuckartem v Německu. Konečně Sinitsin v roce 1914 při svých experimentech s králíky popsal způsob migrace motolic ze střev do jater definitivního hostitele.
Vývojový cyklus a morfologie
Vajíčko
Dospělé motolice se lokalizují v jaterních žlučovodech, kde se živí krví. Po dosažení pohlavní dospělosti kladou vajíčka. Ta postupně odcházejí společně se žlučí přes žlučovody do střeva. S výkaly hostitele se poté vajíčka dostávají do vnějšího prostředí. Vajíčko F. hepatica je oválného tvaru, žlutohnědé barvy, o délce 130–150 μm a šířce 60–90 μm, na jednom pólu opatřené víčkem (operculum). Obsah vajíčka je vyplněn četnými žloutkovými granulemi. S deštěm jsou vajíčka uvolněna z trusu do vodního prostředí, kde se dále vyvíjejí (bez dostatku vody vajíčka hynou). Uvnitř vajíčka se postupně formuje larva, zvaná miracidium. Vývin miracidií uvnitř vajíček, jakož i samotné líhnutí z vajíček, je ovlivněno především teplotou, vlhkostí a tenzí kyslíku. Obecně miracidia se vyvíjejí ve vajíčcích v teplotním rozmezí 10–30 °C s teplotním optimem mezi 23–26 °C. Při teplotách 0–10 °C je vývin vajíčka zastaven, ale vajíčko zůstavá životaschopné (až 2 roky). Při zvýšení teploty nad 10 °C se opět začne vyvíjet miracidium.
Miracidium
Za přirozených podmínek se miracidium vyvine zhruba za 10–20 dní. Jakmile dokončí svůj vývoj, opouští vajíčko odklopením operkula skrze horní pól vajíčka. Vylíhlé miracidium je kyjovitého tvaru, s výraznými očními skvrnami v přední části, měří okolo 130 μm. Povrch miracidia tvoří obrvená kutikula. Vylíhlá miracidia ihned aktivně plavou rychlostí zhruba 1 mm.s−1 a vyhledávají vhodného plže. Miracidia nepřijímají žádnou potravu, jejich energetické výdaje potřebné k pohybu jsou pokryty ze zásobního glykogenu pocházajícího jěště z vajíčka. Miracidia jsou tudíž schopna plavat ve vodě pouze po dobu 12–24 hodin. Larvy, které v tomto čase nestačí proniknout do vhodného mezihostitele, rychle hynou. Chování miracidií většiny druhů motolic je obecně charakterizováno třemi vlastnostmi: pozitivní fototaxe (pohybují se za světlem), negativní geotaxe (tzn. mají tendenci plavat k hladině) a zejména pak pozitivní chemotaxe (jsou přitahovány některými chemickými látkami). Při hledání plže se uplatňuje především posledně jmenovaná vlastnost, kdy miracidia jsou „vábena“ mukopolysacharidy obsaženými ve slizu vodních plžů. Při přichycení k plži se miracidia zavrtají do plášťové dutiny pomocí rostra a sekrece proteáz. Při průchodu do těla plže ztrácí brvy a přeměnují se ve vakovitý útvar zvaný sporocysta.
Sporocysta
Mladá sporocysta migruje skrze lymfatický a krevní oběh plže do hepatopankreatu plže a zde začíná růst. Uvnitř sporocysty se ze zárodečných buněk postupně formují mateřské redie.
Redie
Mateřské redie opouští tělo sporocysty, která tak zaniká. Toto stádium se již živí tkáňovými tekutinami plže, roste a značně narušuje tkáně mezihostitele. Mají vyvinutou travicí trubici začínající ústy, na něž navazuje svalnatý pharynx. Uvnitř každé redie se ze zárodečných buněk formují další generace redií – tzv. dceřiné redie. Někdy se používá místo označení mateřská a dceřiná redie také redie I. a II. genarace. Ve dceřiných rediích (II. generace redií) se již formují ocáskaté larvy – cerkarie. Výzkumy z Francie potvrdily, že se v plži mohou tvořit i III. generace redií.
Cerkarie
Jakmile cerkarie dospějí, aktivně se uvolňují z lumen redie a následně opouštějí i plže. Uvolňování cerkarii (tzv. cercarial shedding) infikovaný plž zpravidla nepřežije a hyne. Cerkarie se skládá z kulaté, mírně srdcovité hlavičky (vlastní tělo) a bičíkatého ocásku, jenž slouží k pohybu ve vodě. Vlastní tělo je vybaveno dvěma přísavkami a typickým vidličkovitě rozvětveným slepě zakončeným střevem. Morfologicky se tak značně podobá dospělé motolici. Hlavička měří 200 μm, ocásek okolo 0,5–0,7 mm. Cerkarie plavou pomocí ocásku ve vodě a ulpívají na rostlinách. Jakmile se cerkarie přichytí k rostlinném porostu, odhazuje bičík a vytváří kolem celého těla ochranný obal (proces zvaný encystace) – vzniká metacerkarie. Celý vývoj larválních stadií v plži trvá 1–2 měsíce a je ovlivněn řadou faktorů jako je teplota, velikost plže, imunitní reakce plže apod. Počty cerkarií z jednoho plže infikovaného jedním miracidiem se liší dle různých autorů. Zatímco Krull uvádí, že z jednoho miracidia F. hepatica může v plži vzniknout až 4000 cerkarií[10], francouzští autoři při svých experimentálních infekcích G. truncatula udávají v průměru 344 cerkarií.
Metacerkarie
Toto infekční a zároveň poslední larvální stádium motolic je pevně přichyceno na povrch rostlin nebo plave na vodní hladině. Jeho metabolismus je omezen na minimum a v podstatě čeká na pozření definitivním hostitelem. Ochranný obal chrání před nepříznivými vlivy. Životaschopnost, respektive infekceschopnost, metacerkarií je ve vlhkém prostředí až několik měsíců. Jsou-li chráněny před mrazem pod sněhovou pokrývkou, jsou schopny přežít i zimní měsíce do dalšího jara.
Dospělá motolice
Definitivní hostitel se nakazí při pastvě konzumací porostu s metacerkariemi, případně vodou. Obal metacerkarie je při průchodu slezem (žaludkem u monogastrů) narušen trávicími enyzmy a začíná tak proces excystace (odhození ochranného obalu), který bývá dokončen v tenkém střevu. Vzniká mladá motolička, jež ihned migruje skrze stěnu střevní do břišní dutiny a dále do jater. Přestože mají mladé motolice F. hepatica silnou afinitu k jaterní tkáni, mohou někeří jedinci domigrovat i do jiných orgánů (ledviny, plíce). Tito však nedosáhnou pohlavní dopělosti a hynou. Jakmile se juvenilní motolice dostanou do jater, migrují skrze jaterní parenchym a rostou. Po průniku do jaterních žlučovodů končí migrace, pohlavně dozrávají a začínají produkovat vajíčka. Doba od průniku metacerkárie do těla hostitele do přítomosti prvních vajíček v trusu (tzv. prepatentní perioda) se pohybuje od 8 do 10 týdnů, výjimečně i 6. Motolice jsou schopny žít ve žlučovodech a produkovat vajíčka až několik let.
Rozšíření
Infekce lidí a zvířat motolicí jaterní byly zaznemenány z celkem pěti kontinentů, druh F. hepatica je tedy rozšířen po celém světě krom Antarktidy. Naproti tomu, areál výskytu blízce příbuzného druhu Fasciola gigantica (rovněž způsobující fasciolózu) je omezen na subtropické a tropické oblasti Afriky a Asie. V některých oblastech Afriky a Asie se tak vyskytují oba druhy současně.
Z mnoha fylogenetických a paleontologických studí vyplývá, že motolice jaterní pochází původně z Evropy. Přítomnost vajíček F. hepatica byla prokázána v koprolitech lidí a zvířat pocházejících z doby kamenné z období pozdního mezolitu (zhruba před 5000-5100 lety) a neolitu, tedy z období rozvoje zemědělství a počátků domestikace zvířat. Nálezy pocházejí z archeologických lokalit v Francii a Švýcarsku. Vajíčka motolice byla rovněž identifikována i z jiných lokalit v Evropě a to jak z doby bronzové, tak např. z období starověkého Říma či ze středověku. Nikdy však nebyla nalezena v prehistorických sedimentech či koprolitech v Americe či Austrálii. F. hepatica byla tedy introdukována do Ameriky, Austrálie a na Nový Zéland a Afriky. V nových oblastech se motolice adaptovala na nové definitivní hostitele (např. lamy a alpaky v Jižní Americe) i mezihostitele (Austropeplea (Lymnaea) tomentosa jako zástupce plovatkovitých v Austrálii).
Mezihostitelé
Jako mezihostitelé F. hepatica figurují vodní plži z čeledi plovatkovitých (Lymnaeidae). Původním a zároveň hlavním mezihostitelem v Evropě je Galba truncatula (bahnatka malá). S rozšířením na další kontinenty se motolice adaptovala i na další plovatkovité plže. V Austrálii je mezihostitelem Austropeplea (Lymnaea) tomentosa, v Severní Americe Pseudosuccinea columella a Fossaria (Galba) bulimoides, v Jižní Americe krom introdukované bahnatky malé také Fossaria (Lymnaea) viatrix, na Kubě a v Karibské oblasti pak Fossaria (Lymnaea) cubensis. Mimo tyto druhy je vnímavá k infekci řada plovatkovitých plžů, kteří již nemají takový význam.
Definitivní hostitelé
Definitivním hostitelem motolice jaterní jsou přežvýkavci, nicméně k infekci jsou vnímaví i další savci, včetně člověka. Mezi hlavní a nejdůležitější hostitele však patří skot a ovce, u nichž figuruje F. hepatica jako jeden z nejvýznamnějších parazitů vůbec. U ostatních druhů nemá motolice jaterní již tak značný ekonomický dopad. Ve volné přírodě je parazit běžný u jelenovitých (v Evropě např. jelen lesní, srnec obecný), volně žijících turovitých (např. muflon, kamzík), ale i dalších nepřežvýkavých druhů jako je ondatra, zajíc, králík apod.